1․Ներկայացնել մեյոզի փուլերը մանրամասն։
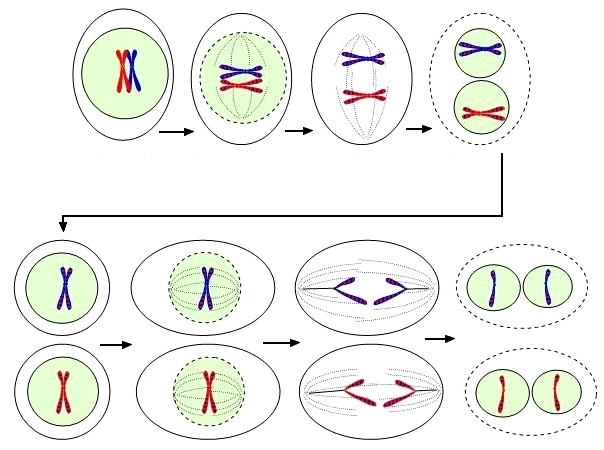
Մեյոզը բաղկացած է 2 հաջորդական բաժանումներից, որոնց միջև կա կարճ ինտերֆազ։
- I պրոֆազ,, առաջին պրոֆազը շատ բարդ է և կազմված է 5 փուլերից։
- Լիպտոտենա կամ լիպտոնեմա, քրոմոսոմների փաթեթավորում, ԴՆԹ-ի կոնդենսացիա քրոմոսոմների ձևավորումով՝ բարակ թելերի տեսքով (քրոմոսոները կարճանում են)։
- Զիգետգենա կամ զիգոնեմա, ընթանում է կոնյուգացիա՝ հոմոլոգ քրոմոսոմների ձևավորված կազմությունների հետ միացումով, որը կազմված է երկու միացած քրոմոսոմներից, որոնք նաև կոչվում են բիվալենտեներ և ընթանում է նրանց հետագա խտացումը։
- Պահիտենա կամ պահինեմա, (ամենաերկար փուլը), մի քանի մասերում հոմոլոգ քրոմոսոմները իրար են միացվում, ձևավորելով խիազմաներ։ Այնտեղ տեղի է ունենում տրամախաչում՝ հոմոլոգիական քրոմոսոմների միջև մասերի փոխանակում։
- Դիպլոտենա կամ դիպլոնեմա, տեղի է ունենում քրոմոսոմների մասնակի ապապարուրում, այդ ժամանակ կարող է նաև աշխատել գենոմի մասը, տեղի է ունենում տրանսկրիպցիաների գործընթացներ (ՌՆԹ-ի ձևավորում), տրանսլիացիա (սպիտակուցի սինթեզ); հոմոլոգ քրոմոսոմները դեռևս մնում են միացված։ Որոշ կենդանիների մոտ ձվաբջիջիների քրոմոսոմները մեյոզի այս փուլում ձեռք են բերում լամպաձև խոզանակի նման քրոմոսոմների բնորոշ ձև։
- Դիակենես, ԴՆԹ-ն մաքսիմալ կերպով կոնդեսացվում է, սինթեզման գործընթացները ավարտվում են, միջուկային թաղանքը լուծվում է; ցենտրիոլները հեռանում են դեպի տարբեր բևեռներ; հոմոլոգիական քրոմոսոմները մնում են միացված։
I պրոֆազից հետո ցենտրիոլները հեռանում են դեպի բջջի բևեռները, ձևավորվում են բաժանման իլիկի թելիկները, միջուկային մեմբրանը և միջուկները քանդվում են։
- I մետաֆազ, բիվալենտային քրոմոսոմները ձևավորվում են բջջի հասարակածի երկայնությամբ։
- I անաֆազ, միկրոխողովակները կրճատվում են, բիվալենտները բաժանվում են և քրոմոսոմները հեռանում են դեպի բևեռները։ Կարևոր է նշել, որ քրոմոսոմների կոնյուգացիայից զիտոգենում դեպի բրեռներ են շարժվում ամբողջական քրոմոսոմներ, որոնցից յուրաքանչյուրը բաղկացած են երկու քրոմատիդներից, այլ ոչ թե առանձին քրոմատիդներից, ինչպես միտոզում։
- I թելոֆազ, քրոմոսոմները ապապարուրվում են և հայտնվում է միջուկային թաղանք։
Մեյոզի երկրորդ բաժանումը տեղի է ունենում առաջինից անմիջապես հետո, առանց ինտերֆազի արտահայտման. S շրջանը բացակայում է, քանի որ երկրորդից առաջ տեղի չի ունենում ԴՆԹ-ի կրկնապատկում։
- II պրոֆազ, տեղի է ունենում քրոմոսոմների կոնդեսացիա, բջջակենտրոնը բաժանվում է և բաժանման մասերը շարժվում են դեպի բևեռներ, քանդվում է միջուկային թաղանթը, ձևավորվում է իլիկի թելիկներ, որն ուղղահայաց է առաջին բաժանմանը։
- II մետաֆազ, ունիվալենտային քրոմոսոմները (որոնցից յուրաքանչյուրը բաղկացած է երկու քրոմատիդներից) մակերևույթներից վրա տեղավորվում են «էկվատորում», ձևավորելով այսպես կոչված մետաֆազային շերտ։
- II անաֆազ, ունիվալենտները բաժանվում են և քրոմատիդները շարժվում են դեպի բևեռներ։
- II թելոֆազ, քրոմոսոմները ապապարուրվում են և հայտնվում է միջուկային շերտ։
2․Որ բջիջներն են բազմանում մեյոզի ճանապարհով։
3․Ինչով են իրարից տարբերվում մեյոզը և միթոզը։